Central dogma of molecular biology

This article is part of the series on: Gene expression |
|||
| Introduction to Genetics | |||
| General flow: DNA > RNA > Protein | |||
| special transfers (RNA > RNA, RNA > DNA, Protein > Protein) |
|||
| Genetic code | |||
| Transcription | |||
| Transcription (Transcription factors, RNA Polymerase,promoter) Prokaryotic / Archaeal / Eukaryotic |
|||
| post-transcriptional modification (hnRNA,5' capping,Splicing,Polyadenylation) |
|||
| Translation | |||
| Translation (Ribosome,tRNA)
Prokaryotic / Archaeal / Eukaryotic |
|||
| post-translational modification (functional groups, peptides, structural changes) |
|||
| gene regulation | |||
| epigenetic regulation (Genomic imprinting) |
|||
| transcriptional regulation | |||
| post-transcriptional regulation (sequestration, alternative splicing,miRNA) |
|||
| translational regulation | |||
| post-translational regulation (reversible,irreversible) |
|||

The central dogma of molecular biology was first articulated by Francis Crick in 1958[1] and re-stated in a Nature paper published in 1970:[2]
- The central dogma of molecular biology deals with the detailed residue-by-residue transfer of sequential information. It states that information cannot be transferred back from protein to either protein or nucleic acid.
In other words, 'once information gets into protein, it can't flow back to nucleic acid.'
The dogma is a framework for understanding the transfer of sequence information between sequential information-carrying biopolymers, in the most common or general case, in living organisms. There are 3 major classes of such biopolymers: DNA and RNA (both nucleic acids), and protein. There are 3×3 = 9 conceivable direct transfers of information that can occur between these. The dogma classes these into 3 groups of 3: 3 general transfers (believed to occur normally in most cells), 3 special transfers (known to occur, but only under specific conditions in case of some viruses or in a laboratory), and 3 unknown transfers (believed never to occur). The general transfers describe the normal flow of biological information: DNA can be copied to DNA (DNA replication), DNA information can be copied into mRNA, (transcription), and proteins can be synthesized using the information in mRNA as a template (translation).[2]
Contents |
Biological sequence information
The biopolymers DNA, RNA and proteins, are linear polymers (i.e.: each monomer is connected to at most two other monomers). The sequence of their monomers effectively encodes information. The transfers of information described by the central dogma are faithful, deterministic transfers, wherein one biopolymer's sequence is used as a template for the construction of another biopolymer with a sequence that is entirely dependent on the original biopolymer's sequence.
General transfers of biological sequential information
-
Table of the 3 classes of information transfer suggested by the dogma General Special Unknown DNA → DNA RNA → DNA protein → DNA DNA → RNA RNA → RNA protein → RNA RNA → protein DNA → protein protein → protein
DNA Replication
As the final step in the Central Dogma, to transmit the genetic information between parents and progeny, the DNA must be replicated faithfully. Replication is carried out by a complex group of proteins that unwind the superhelix, unwind the double-stranded DNA helix, and, using DNA polymerase and its associated proteins, copy or replicate the master template itself so the cycle can repeat DNA → RNA → protein in a new generation of cells or organisms.
Transcription
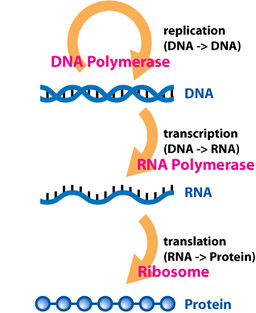
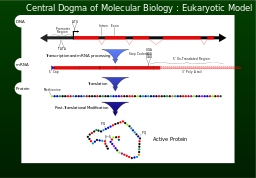
Transcription is the process by which the information contained in a section of DNA is transferred to a newly assembled piece of messenger RNA (mRNA). It is facilitated by RNA polymerase and transcription factors. In eukaryote cells the primary transcript (pre-mRNA) is often processed further via alternative splicing. In this process, blocks of mRNA are cut out and rearranged, to produce different arrangements of the original sequence.
Translation
Eventually, this mature mRNA finds its way to a ribosome, where it is translated. In prokaryotic cells, which have no nuclear compartment, the process of transcription and translation may be linked together. In eukaryotic cells, the site of transcription (the cell nucleus) is usually separated from the site of translation (the cytoplasm), so the mRNA must be transported out of the nucleus into the cytoplasm, where it can be bound by ribosomes. The mRNA is read by the ribosome as triplet codons, usually beginning with an AUG, or initiator methionine codon downstream of the ribosome binding site. Complexes of initiation factors and elongation factors bring aminoacylated transfer RNAs (tRNAs) into the ribosome-mRNA complex, matching the codon in the mRNA to the anti-codon in the tRNA, thereby adding the correct amino acid in the sequence encoding the gene. As the amino acids are linked into the growing peptide chain, they begin folding into the correct conformation. This folding continues until the nascent polypeptide chains are released from the ribosome as a mature protein. In some cases the new polypeptide chain requires additional processing to make a mature protein. The correct folding process is quite complex and may require other proteins, called chaperone proteins. Occasionally, proteins themselves can be further spliced; when this happens, the inside "discarded" section is known as an intein.
Special transfers of biological sequential information
Reverse transcription

Reverse transcription is the transfer of information from RNA to DNA (the reverse of normal transcription). This is known to occur in the case of retroviruses, such as HIV, as well as in eukaryotes, in the case of retrotransposons and telomere synthesis.
RNA replication
RNA replication is the copying of one RNA to another. Many viruses replicate this way. The enzymes that copy RNA to new RNA, called RNA-dependent RNA polymerases, are also found in many eukaryotes where they are involved in RNA silencing.[3]
Direct translation from DNA to protein
Direct translation from DNA to protein has been demonstrated in a cell-free system (i.e. in a test tube), using extracts from E. coli that contained ribosomes, but not intact cells. These cell fragments could express proteins from foreign DNA templates, and neomycin was found to enhance this effect.[4][5]
Methylation
Variation in methylation states of DNA can alter gene expression levels significantly. Methylation variation usually occurs through the action of DNA methylases. When the change is heritable, it is considered epigenetic. When the change in information status is not heritable, it would be a somatic epitype. The effective information content has been changed by means of the actions of a protein or proteins on DNA, but the primary DNA sequence is not altered.
Prions - almost a "known transfer"
Prions are proteins that propagate themselves by making conformational changes in other molecules of the same type of protein. This change affects the behaviour of the protein. In fungi this change happens from one generation to the next, i.e. Protein → Protein. While this represents a transfer of information, prion interactions leave the sequence of the protein unchanged, and so are not technically considered an exception to the central dogma. This is based upon a narrow interpretation of the central dogma and its role in evolutionary processes. The conclusion that the amino acid sequence of a protein is the ultimate arbiter of protein conformation(see Anfinsen's Dogma), an idea also violated by prions as well as conformational change induced by silent mutations in the MDR1 gene, is likely to continue to be undermined as new tools are developed to observe the cellular diversity of protein conformations. Sequences, are not directly selected for by evolutionary processes. Protein conformations, and their resulting functions and traits are subject to selection. A "technical exception" which demonstrates evolutionary principles is not an exception.
As of 2009, prion replication has been shown to be subject to mutation and natural selection similar to other forms of replication.[6]
Use of the term "dogma"
In his autobiography, What Mad Pursuit, Crick wrote about his choice of the word dogma and some of the problems it caused him:
I called this idea the central dogma, for two reasons, I suspect. I had already used the obvious word hypothesis in the sequence hypothesis, and in addition I wanted to suggest that this new assumption was more central and more powerful. ... As it turned out, the use of the word dogma caused almost more trouble than it was worth.... Many years later Jacques Monod pointed out to me that I did not appear to understand the correct use of the word dogma, which is a belief that cannot be doubted. I did apprehend this in a vague sort of way but since I thought that all religious beliefs were without foundation, I used the word the way I myself thought about it, not as most of the world does, and simply applied it to a grand hypothesis that, however plausible, had little direct experimental support.
Similarly, Horace Freeland Judson records in The Eighth Day of Creation:[7]
"My mind was, that a dogma was an idea for which there was no reasonable evidence. You see?!" And Crick gave a roar of delight. "I just didn't know what dogma meant. And I could just as well have called it the 'Central Hypothesis,' or — you know. Which is what I meant to say. Dogma was just a catch phrase."
See also
- Weismann barrier
- Genetic code
- Alternative splicing
- Riboswitch
|
||||||||||||||||||||||||||||||
References
- ↑ Crick, F.H.C. (1958): On Protein Synthesis. Symp. Soc. Exp. Biol. XII, 139-163. (pdf, early draft of original article)
- ↑ 2.0 2.1 Crick, F. (1970): Central Dogma of Molecular Biology. Nature 227, 561-563. PMID 4913914
- ↑ Ahlquist P (May 2002). "RNA-dependent RNA polymerases, viruses, and RNA silencing". Science (journal) 296 (5571): 1270–3. doi:10.1126/science.1069132. PMID 12016304.
- ↑ B. J. McCarthy and J. J. Holland (September 15, 1965). "Denatured DNA as a Direct Template for in vitro Protein Synthesis". Proceedings of the National Academy of Sciences of the United States 54: 880–886. doi:10.1073/pnas.54.3.880. PMID 4955657.
- ↑ .T. Uzawa, A. Yamagishi, T. Oshima (2002-04-09). "Polypeptide Synthesis Directed by DNA as a Messenger in Cell-Free Polypeptide Synthesis by Extreme Thermophiles, Thermus thermophilus HB27 and Sulfolobus tokodaii Strain 7". The Journal of Biochemistry 131: 849–853. PMID 12038981.
- ↑ Li J, Browning S, Mahal SP, Oelschlegel AM, Weissmann C (December 2009). "Darwinian Evolution of Prions in Cell Culture". Science. doi:10.1126/science.1183218. PMID 20044542. Lay summary.
- ↑ Horace Freeland Judson (1996). "Chapter 6: My mind was, that a dogma was an idea for which there was no reasonable evidence. You see?!". The Eighth Day of Creation: Makers of the Revolution in Biology (25th anniversary edition). Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press. ISBN 0-87969-477-7.